In this article we will discuss about carbon dioxide fixation during photosynthesis in crop plants.
Carbon Dioxide Fixation – C3 Species:
The path of carbon in photosynthesis, which forms the basis of our present knowledge, has been worked out by Calvin and co-workers. This Calvin cycle is outlined in Figure 1.13. The CO2 fixation portion of the Calvin cycle is catalyzed by the enzyme ribulose bis-phosphate (RuBP) carboxylase. The ATP produced in photophosphorylation is used to convert ribulose-5-phosphate to RuBP (Fig. 1.13).

After CO2 fixation, ATP along with reduced nucleotides from the light process, change 3-phosphoglyceric acid (3-PGA) to 3- phosphoglyceraldehyde (3-PGald). Species with this pathway are called C3 pathway species because the first product that can be measured after adding radioactive CO2 (14CO2) is a three-carbon molecule, 3-PGA.
Carbon Dioxide Fixation-C4 Species:
From 1954 to 1966 the Calvin cycle was considered the only pathway for CO2 fixation in higher plants. Then Hatch and Slack (1966), working in Australia, presented detailed evidence that another pathway for CO2 fixation exists in some species. This pathway incorporates CO2 using phosphoenol pyruvate (PEP) carboxylase enzyme. The ATP produced in photophosphorylation is used to convert pyruvate to PEP.
The PEP, a three-carbon molecule, is carboxylated to three four-carbon acids (oxaloacetate, malate, and aspartate). These acids are translocated to vascular sheath cells where they are converted to pyruvate.
In the change to pyruvate, a carbon is released that is converted, either by addition to RuBP or by addition to a two-carbon molecule, to 3-PGA by RuBP carboxylase. After 3-PGA is produced, the Calvin cycle is operative. Species with the Hatch and Slack pathway are called C4 species because the first product of photosynthesis in the mesophyll is a four-carbon molecule.
Comparing C3 and C4 Species of Carbon Dioxide Fixation:
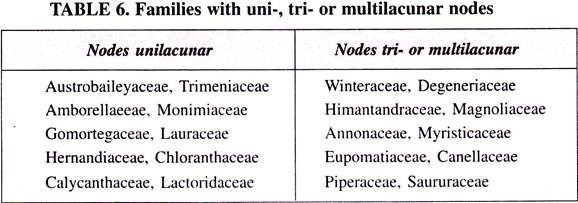
Comparisons between the species using these two pathways show many differences:
1. Anatomical differences (The leaf anatomy of C4 species is called the Kranz anatomy).
a. The C4 species have chloroplasts in the vascular sheath cells; C3 species do not.
b. Chloroplasts in the mesophyll of C3 and C4 species look similar (they usually have double external membranes and well-developed grana) but are very different biochemically. In C3 species, CO2 is fixed by RuBP carboxylase, the Calvin cycle is operative, and starch is accumulated (Fig. 1.13). In C4 species, CO2 is fixed by PEP carboxylase, which forms four-carbon acids that translocate to vascular sheath cells. Starch is not formed in these mesophyll cells, only four-carbon acids.
c. Chloroplasts in vascular sheath cells of C4 species are anatomically different. They are larger and have less-developed grana than in mesophyll cell chloroplasts; and since the Calvin cycle is operative, they store starch.
2. PEP carboxylase enzyme has a greater affinity for CO2 than does the RuBP carboxylase enzyme, so it can operate more efficiently at low CO2 concentrations.
3. Species with the C4 pathway generally have higher photosynthetic rates than do C3 species, especially at high light intensities.
4. C4 species may use more energy than C3 species to fix one CO2 molecule. This point has not been proven but seems probable, as ATP is required to form PEP.
5. Ribulose bis-phosphate carboxylase is present at much lower levels in C4 species than in C3 species (i.e., around 10%). C3 species apparently do not have PEP carboxylase present.
6. Differences in adaptation occur for species with different CO2 fixation mechanisms: C3 species are adapted to cool and moist to hot and moist conditions while C4 species are adapted to hot, dry, or moist conditions.
7. The main factor causing the increased photosynthetic efficiency of C4 species is the lack of measurable photorespiration (respiration in the light). It results in CO2 loss in photosynthetic tissue and is a major source of CO2 evolution by C3 species in the light. It occurs as a by-product of the Calvin cycle; since RuBP carboxylase is also RuBP oxygenase, the O2 and CO2 compete for the same enzyme and for the same ribulose bis- phosphate substrate.
Photorespiration is not significant in C4 species; this is believed to be the major factor giving C4 species higher photosynthetic efficiency than C3 species. The C4 species are believed to have little or no photorespiration because movement of four-carbon acids into the vascular sheath cells concentrates CO2 in these cells, which would favor the RuBP carboxylase reaction over RuBP oxygenase.
However, any CO2 evolved from the vascular sheath cells would probably not leave the leaf because of the great affinity of PEP carboxylase for CO2 in mesophyll cells; thus any photorespiration that did occur could not be measured. Photorespiration evolves CO2 without seemingly coupling the energy acquired to a useful purpose. Photorespiration does, however, provide amination for amino acid synthesis and keeps cycling inorganic phosphate, which may be beneficial under low light and cool temperatures.
Special techniques must be used to measure photorespiration:
a. Air without CO2 is passed over an illuminated leaf. If CO2 is evolved, it is a measure of photorespiration.
b. Plants or leaves in a gas-tight container under illumination will pull down the CO2 concentration in the air until it comes to an equilibrium (compensation concentration), which is a measure of photorespiration.
c. If a leaf is suddenly put into darkness, photosynthesis will stop but photorespiration will continue a short while to use up the glycolic acid. This causes a post-illumination burst of CO2 evolution much greater than the equilibrium of CO2 evolution of dark respiration.
d. Oxygen (O2) is required for the conversion of glycolic acid to glyoxylic acid. If O2 is reduced in the air from 21 to 1% or less, the photorespiration is stopped. Therefore, the difference in photosynthesis at O2 concentrations of 21 and 1% is a measure of photorespiration.
Both types of CO2 fixation occur in crop species.
Crassulaceous Acid Metabolism for CO2 Fixation:
A third type of CO2 fixation, called crassulaceous acid metabolism (CAM), occurs primarily in succulent plants, which have fleshy leaves or stems. Such plants are adapted to arid conditions where low transpiration (evaporation from plant surfaces) is a survival necessity.
Under low moisture conditions they open their stomata at night to absorb CO2 and close them in the day, thus reducing the transpiration load of the plant. There are only a few CAM plants classified as domestic crops; these include pineapple, Agave (sisal, henequen. and others), and prickly pear.
The CAM species fix CO2 into four-carbon acids with PEP carboxylase as do the C4 species, only it occurs at night when stomata are open and the energy required comes from glycolysis.
Solar radiation causes stomatal closure and irradiates the leaf; this light energy is used to drive the Calvin cycle, taking CO2 from the four-carbon acids as in the reaction in the bundle sheath cells of C4 species. The chloroplasts of CAM plants are more similar to those of the C3 species. Under favorable moisture conditions, many CAM species, change stomatal function and carboxylation is similar to that of C3 species.
Thus, CAM plants have developed an ingenious physiological method of reducing moisture loss and escaping drought. They often are important crop plants where moisture availability for crops remains low.
Comments are closed.