In this article we will discuss about the anatomy of flowering plants.
Plant anatomy deals with the study of internal structures of various parts of the plant body like root, stem and leaf. All these organs are made up of different tissues.
The Tissues:
A vascular plant begins its existence as a morphologically simple unicellular zygote. The zygote develops into an embryo and thereafter into a mature sporophyte. The development of the mature sporophyte involves organisation of cells into tissues and tissue systems.
Histology is the study of tissues. A tissue may be defined as a group cells that perform a definite function and have a common origin.
Plant tissues can be broadly classified as follows:
1. Meristematic tissues.
2. Permanent tissues.
1. Meristematic Tissues: (Gr., meristos = divisible):
A meristematic tissue is an undifferentiated mass of cells that are in a continuous state of division or retain their power of division.
The characteristic features of meristematic tissues are:
i. They are living and consist of an undifferentiated mass of rapidly dividing cells.
ii. Cells are spherical, polygonal or rectangular in shape.
iii. They are compactly arranged without intercellular spaces.
iv. Cells are interconnected by plasmodesmata.
v. Cell wall is thin, with only a primary wall made up of cellulose. Secondary wall is absent.
vi. Cytoplasm is dense. Vacuoles are small or completely absent.
vii. Nucleus is either in interphase or in divisional stages.
viii. Plastids occur as proplastids. Cells do not store reserve food material. Ergastic substances are absent.
ix. Meristematic tissues are found in the growing regions of the plant body and contribute to plant growth.
Functions of Meristem:
i. They are responsible for growth in the plant body by adding new cells followed by differentiation. These differentiated cells form permanent tissues.
ii. They are responsible for the formation of new organs like buds, flowers, leaves etc.
iii. They help to heal the injured parts of the plant body.
2. Permanent Tissues:
Permanent tissues are derived from meristems. They are composed of cells that have lost the power of cell division and have attained a definite form and size. Under certain circumstances, they may regain the power of cell division. They may be living or dead on maturity.
Permanent tissues are classified into two groups:
I. Simple permanent tissues.
II. Complex permanent tissues.
The Tissue Systems:
The different plant tissues irrespective of their position in the plant body performing a common function constitute the tissue system. The tissues in a system may have no structural similarity and may also differ in their origin.
Sachs (1875) distinguished three tissue systems in plants:
A. Epidermal tissue system.
B. Ground or fundamental tissue system.
C. Vascular tissue system.
A. Epidermal Tissue System:
(a) Epidermis:
It forms outer covering of plant organs which is in direct contact with the external environment. The word epidermis is derived from two greek words ‘epi’ – means upon and ‘derma’ means skin. Generally it is associated with structures like stomata and hairs (trichomes). The epidermis of roots is called epiblema or piliferous layer.
Epidermis is generally made up of single layered living, parenchymatous cells, which are compactly arranged forming a continuous layer (interrupted by stomata in leaves). The outer walls of epidermis are usually thicker when compared to the inner. Externally the epidermis is lined by a layer of cuticle. Sometimes a waxy coating may be present. These help to minimise transpiration.
Epidermal cells have a large central vacuole and peripheral thin cytoplasm. In some cases epidermis is many layered – multiple epidermises, E.g. – Ficus (Moraceae), Nerium (Apocynaceae). In epiphytic roots of orchids, e.g. – Vanda, the multiple epidermises is modified into velamen which can absorb water (moisture) from the atmosphere.
Some cells of velamen are non-living with spiral or reticulate or pitted thickenings. In some xerophytic grasses, some of the epidermal cells become larger, thin walled and are vacuolated. These cells are called bulliform cells or motor cells. In dry conditions, these cells lose water and help in the rolling of leaves and thus reduce water loss by transpiration.
(b) Stomata:
Stomata are microscopic openings present in the epidermis of leaves and young stem. Each stoma is a slit like opening bound by two specialised epidermal cells called guard cells on either side. The guard cells are bean/kidney shaped in dicots and dumb-bell shaped in monocots. The guard cells are surrounded by a few epidermal cells called subsidiary cells.
The guard cells have thick walls towards the stomatal aperture and thin walls on the outerside. The cellulose micelles in guard cells are oriented radially. Further, guard cells have special elastic properties. These help them to stretch and contract during the opening and closing mechanism of Stoma. The protoplasm of guard cells contains chloroplasts.
The number of stomata per sq. mm of leaf is 14 to 1000, most generally it is 50-300.
In dicots, five main types of stomata are distinguished:
(a) Anomocytic (Rannunculus Type):
Number of subsidiary cells is five.
(b) Anisocytic (Cruciferous Type):
Number of subsidiary cells three and one is smaller.
(c) Paracytic (Rubiaceous Type):
Number of subsidiary cells is two and these two are parallel to the long axis of guard cells.
(d) Diacytic (Caryophyllaceous Type):
Number of subsidiary cells is two and these are at right angles to guard cells.
When stomata are mostly found on the lower epidermis, such leaves are called hypostomatic. When they occur on the upper epidermis, they are called epistomatic. When stomata occur on both surfaces, the leaves are called amphistomatic.
Based on their distribution, they are of five types:
I. Apple Type:
Stomata only on the underside of a leaf. E.g. – apple, mulberry.
II. Potato Type:
More stomata on the undersurface. E.g. – potato, cabbage, bean.
III. Oat Type:
Stomata almost equal on the two surfaces. E.g. – maize, oat.
IV. Water Lily Type:
Stomata more on the upper surface. E.g. – many aquatic plants.
V. Potamogeton Type:
Stomata vestigeal or absent. E.g. – Potamogeton.
In xerophytes, the number of stomata is reduced. In many xerophytes, stomata are sunken to reduce water loss by transpiration.
(e) Trichomes:
Trichomes are epidermal hairy outgrowths present permanently or temporarily. They may be unicellular or multicellular.
They are of following types:
(a) Unicellular hairs. E.g. – Cannabis.
(b) Multicellular hairs. E.g. – Tridax.
(c) Stellate hairs. E.g. – Solanum.
(d) Glandular hairs (colleters). E.g. – shaggy glands of Tabernaemontana.
(e) Scales (peltate trichomes). E.g. – Gossypium.
(f) Root hairs. E.g. – tubular extensions in root epidermis.
Functions of Epidermis:
1. Provides protection.
2. Reduces evaporation of water.
3. Gaseous exchange takes place through stomata.
4. Transpiration takes place through stomata.
5. Glandular trichomes secrete various products.
6. Root hairs help in the absorption of water and minerals.
B. Ground Tissue System:
This tissue system is present between the epidermis and vascular tissue system. This system forms the bulk of body and it consists of parenchyma, collenchyma, sclerenchyma, glandular and laticifer tissues.
The different components of this system are:
(a) Hypodermis:
This is the region situated just internal to the epidermis and as an outer region of the cortex. It consists of one, two or few continuous or discontinuous layers of collenchyma (in dicots) or sclerenchyma (in monocots). It is protective and mechanical in function.
(b) General Cortex:
It is mainly parenchymatous. The cells of cortex contain starch grains, oil, tannins and crystals. Sometimes cortical cells may contain chloroplasts and are called chlorenchyma. In hydrophytes the cortex may be aerenchymatous. Special types of cells like sclereids, resin ducts, oil glands, laticifers are found in this region. The cortex helps in vital functions, storage, etc.
(c) Endodermis:
This represents the innermost layer of the cortex. It is generally made up of a single layer of living cells which are barrel shaped and arranged without intercellular spaces. In stems, it stores starch and is called starch sheath (endodermoid layer). In root endodermis, the radial and tangential walls possess special thickenings called casparian strips (discovered by Caspary) which mainly consist of suberin.
The cells of the endodermis opposite the protoxylem elements lack the deposition of suberin and are called passage cells. The function of endodermis is to control the movement of water between cortex and xylem. It also helps to maintain the root pressure.
(d) Pericycle:
This is a single layer with compactly arranged parenchyma cells present internal to the endodermis. It represents the outermost part of stele. In stems, the pericycle may be multilayered. Pericycle is multilayered sclerenchymatous in Cucurbita stem and it is multilayered with alternating patches of parenchyma and sclerenchyma in Tridax stem. Functionally, in roots pericycle gives rise to lateral roots and in dicot roots vascular cambium and cork cambium originate in the pericycle.
(e) Pith and Medullary Rays:
Pith, also called medulla is the central region of dicot stem, dicot and monocot roots. It is generally made up of parenchyma cells. In some cases the pith is made up of sclerenchyma. Sometimes, as in some hydrophytes, pith becomes a hollow cavity. The main function of pith is storage of water and food. The parenchyma cells between the vascular bundles in a dicot stem are radially elongated and extend from the pith outwards. These constitute the medullary rays (pith rays), which help in radial translocation of organic food and water.
C. The Vascular Tissue System:
This includes the vascular tissues i.e., xylem and phloem. They occur together in the plant organs as vascular bundles. A vascular bundle in addition to xylem and phloem, sometimes has cambium in between. These tissues originate from the procambium and apical meristems. The arrangement of xylem and phloem is characteristic to particular plant organs.
A vascular bundle consists of a strand like portion having xylem and phloem of the primary vascular system.
According to the arrangement of xylem and phloem in the vascular bundles, there are three types of bundles namely radial, conjoint and concentric:
(i) Radial:
Here the xylem and phloem form separate bundles and they lie on different radii, side by side. E.g. – roots of seed plants.
(ii) Conjoint:
Here the xylem and phloem are present together in the same bundle on the same radius. Conjoint bundles are collateral and bicollateral.
(a) Collateral:
The xylem and phloem lie together on the same radius. The xylem lies inwards and the phloem outwards.
Here the phloem occurs on one side of the xylem strand. In a dicot stem, the cambium is found to be present in between the xylem and phloem, such bundles are called open. E.g., Helianthus (sunflower). When the cambium is absent, the vascular bundle is called as a closed bundle. E.g., Zea mays (maize).
(b) Bicollateral:
In some bundles the phloem is found to be present on both sides of the xylem. Simultaneously two cambium strips also occur. The various elements are arranged in the following sequence – Outer phloem, outer cambium, xylem, inner cambium and inner phloem. Such bundles are commonly found in the members of Cucurbitaceae. Such bundles are always open.
(iii) Concentric:
Here the xylem surrounds or ensheaths the phloem or vice-versa. The concentric bundles may be of two sub types, amphivasal and amphicribal. If the xylem surrounds the phloem it is called amphivasal (leptocentric) bundle as found in Dracaena and Yucca. If the phloem surrounds the xylem, it is, called amphicribal, (hadrocentric) bundle as found in many ferns. Such bundles are always closed.
Depending on the position of the protoxylem in the vascular bundles.
The following conditions of the xylem are seen:
I. Endarch:
Protoxylem faces the centre of the plant organ.
II. Exarch:
Protoxylem faces the periphery of the plant organ.
III. Mesarch:
Protoxylem is situated in the centre.
Anatomy of Dicotyledonous and Monocotyledonous Plant Organs:
Dicotyledonous Root:
A transverse section of a young dicot root, E.g. – Cicer (Fig. 6.18) shows differentiation into three regions viz.; outer epiblema, middle cortex and central stele. Epiblema or piliferous layer is made up of single layer of parenchyma. Some of the cells of epiblema produce unicellular root hairs which are absorptive in function. Cortex is multilayered, homogenous, made up of loosely arranged parenchyma. Innermost layer of cortex is called endodermis.

It is made up of single layer of barrel shaped parenchyma which shows casparian thickenings along inner tangential and radial walls. Almost all endodermal cells show casparian thickenings, except the cells which are present opposite to protoxylem. These thin walled endodermal cells found opposite to the protoxylem are called passage cells (do not contain casparian thickenings).
Stele has an outer boundary called pericycle made up of single layer of rectangular compactly arranged parenchyma cells. Stele shows vascular bundles which are radial (xylem and phloem arranged in separate bundles along different radii), tetrarch (vascular bundles are four in number) and exarch (protoxylem found towards periphery and metaxylem towards centre). In between xylem and phloem there is parenchymatous conjunctive tissue which helps in secondary growth. There is a small parenchymatous pith at the centre in young dicot roots which is absent in older roots.
Monocotyledonous Root:
A transverse section of a young monocot root, E.g. – maize (Fig. 6.19) shows differentiation into three regions viz.; outer epiblema, middle cortex and central stele. Epiblema or piliferous layer is made up of single layer of parenchyma. Some of the cells of epiblema produce unicellular root hairs which are absorptive in function. Cortex is multilayered, homogenous, made up of loosely arranged parenchyma.
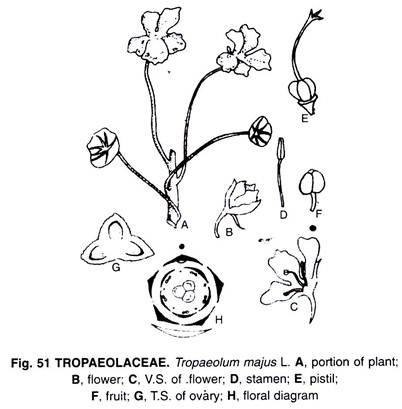
Innermost layer of cortex is called endodermis. It is made up of single layer of barrel shaped parenchyma cells which show casparian thickenings along inner tangential and radial walls. Almost all endodermal cells show casparian thickenings, except the cells which are present opposite to protoxylem. These thin walled endodermal cells found opposite to the protoxylem are called passage cells (do not contain casparian thickenings).
Stele shows an outer boundary called pericycle made up of single layer of rectangular parenchyma cells. Vascular bundles are radial (xylem and phloem arranged in separate bundles along different radii), polyarch (many vascular bundles) and exarch (protoxylem found towards periphery and metaxylem towards centre). In between xylem and phloem there is less amount of parenchymatous conjunctive tissue which does not bring about secondary growth. There is a large parenchymatous pith at the centre.
Dicot Stem:
A transverse section of a young dicot stem, E.g. – Tridax (Fig 6.20) shows outer epidermis, middle cortex and central stele. Epidermis is single layered, parenchymatous and shows multicellular hairs called trichomes. Epidermis is externally lined with a cuticle. It is followed by hypodermis made up of collenchyma.
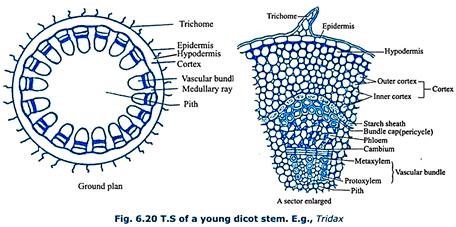
Next to hypodermis, cortex is present, which is divided into outer cortex and inner cortex. Outer cortex is made up of chlorenchyma and inner cortex is made up of parenchyma. Innermost layer of the cortex is called starch sheath (endodermoid layer). It is single layered, parenchymatous and shows deposition of starch grains. It is wavy in outline.
Stele shows an outer boundary called pericycle which is made up of alternating patches of parenchyma and sclerenchyma. Sclerenchymatous portion of the pericycle found above each vascular bundle is called bundle cap. Stele is called eustele. It consists of many vascular bundles arranged in the form of a broken ring around a large pith. Vascular bundles are conjoint, collateral and open with endarch xylem. Cambium is present in between xylem and phloem which helps in secondary growth. Parenchymatous medullary rays are present in between the vascular bundles.
Monocot Stem:
A transverse section of a young monocot stem, E.g. – maize (Fig 6.21) shows an outer single layered epidermis made up of compactly arranged parenchyma cells. Epidermis is lined externally with cuticle. Epidermis is followed by hypodermis which is made up of 2-3 layers of sclerenchyma.
Inner to hypodermis there is an undifferentiated parenchymatous ground tissue. There are many vascular bundles scattered in the ground tissue. This type of stele is described as atactostele. The vascular bundles present towards the periphery are more in number, smaller in size and the ones that are present towards the centre are larger in size but fewer in number.
The vascular bundles are conjoint, collateral and closed. Each vascular bundle consists of xylem and phloem. The arrangement of xylem vessels can be represented by using an alphabet Y. Each arm of Y is represented by metaxylem. The stem of Y is represented by one or two protoxylem elements. The base of Y is represented by the lysigenous cavity. Early formed protoxylem vessels breakdown to form lysigenous cavity or water containing cavity.
Phloem parenchyma is absent. The outermost portion of the phloem which is a broken mass is the protophloem and inner portion is the metaphloem. Protophloem gets disorganised while metaphloem shows sieve tubes and companion cells. Vascular bundles are covered over by sclerenchymatous bundle sheath. Cortex, endodermis, pericycle, medullary rays and pith are not differentiated.
Dorsiventral (Dicotyledonous) Leaf:
A transverse section of a dicot leaf, E.g. – Helianthus (Fig. 6.22) shows an upper and lower epidermis made up of single layer of parenchyma cells that are externally lined with cuticle. Upper epidermis is lined by thick cuticle and lower epidermis is lined by a thin cuticle. Upper epidermis is continuous whereas lower epidermis is discontinuous due to the presence of stomata.
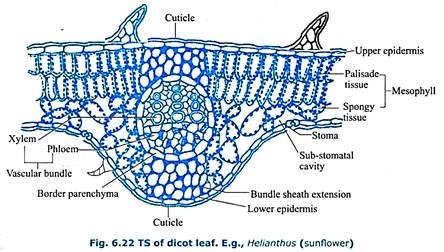
Such a leaf is said to be hypostomatic. Stomata have been shaped guard cells. In between the two epidermal layers there is a mesophyll tissue. It is differentiated into upper palisade parenchyma and lower spongy parenchyma which contain chloroplasts. The vascular bundles are embedded in the mesophyll tissue.
The vascular bundles consist of xylem towards upper epidermis and phloem towards lower epidermis. Cambium is absent (vascular bundles are conjoint, collateral and closed). There is a parenchymatous bundle sheath or border parenchyma around each vascular bundle.
In between the bundle sheath and the epidermal layers there is collenchymatous bundle sheath extension. The dicot leaf shows morphological, anatomical and functional differentiation along dorsal and ventral regions. Therefore, the leaf is said to be dorsiventral leaf.
Isobilateral (Monocotyledonous) Leaf:
A transverse section of a monocot leaf, e.g. – maize (Fig. 6.23) shows an upper and lower epidermis made up of single layer of parenchyma that is externally lined with a cuticle. The amount of cuticle lining the upper epidermis and lower epidermis is same. Both upper as well as lower epidermis is discontinuous due to the presence of stomata.
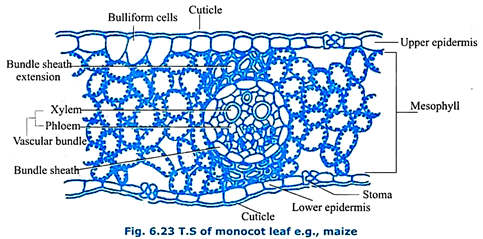
Such a leaf is said to be an amphistomatic leaf. Guard cells are dumb-bell shaped. Upper epidermis shows certain enlarged, hygroscopic cells called bulliform cells or motor cells. They check the rate of transpiration. In between the two epidermal layers there is an undifferentiated mesophyll tissue which is photosynthetic.
The vascular bundles consist of xylem towards upper epidermis and phloem towards lower epidermis. Cambium is absent. There is a parenchymatous bundle sheath or border parenchyma around each vascular bundle which is conjoint, collateral and closed.
In between the bundle sheath and the epidermal layers there is a sclerenchymatous bundle sheath extension. The monocot leaf shows morphological, anatomical and functional similarities along dorsal and ventral regions. Therefore, the leaf is said to be an isobilateral leaf.
Secondary Growth:
Increase in thickness or girth of the stem or root of dicots due to addition of secondary tissues cut off by the vascular cambium and cork-cambium in the stelar and extra-stelar regions respectively is called secondary growth. In perennial dicots (shrubs and trees) after the primary tissues are fully formed, secondary tissues are added to the primary ones and as a result the stem or root increases in thickness.
A. Secondary Growth in a Dicot Stem:
1. Stelar Secondary Growth:
In a dicot plant, secondary growth in the stem occurs both in the stele and in the cortex. This is the secondary growth taking place in the stelar region. The dicot stem, in its primary state of growth contains narrow layers of intrafascicular cambium in between the xylem and phloem.
The parenchyma cells of the primary medullary rays adjacent to the intrafascicular cambium undergo dedifferentiation and give rise to interfascicular cambium. This joins the intrafascicular cambium on either side to form a complete ring of meristem called the cambial ring.
Activity of the Cambial Ring:
This ring is meristematic in nature and it gives off new cells to both its inner and outer sides. The cambial ring is made up of two types of cells-ray initials and fusiform initials. Cells added to the inner side of cambium ring by the division of the fusiform initials gradually become the elements of the secondary xylem where as those cells added to the outer side of the cambium become elements of the secondary phloem. Cells added by the division of the ray initials to the inside as well as outside become elements of the secondary medullary rays. They are one, two or a few layers in thickness, and one to many layers in height.
The cambium is more active on the inner side than on the outer. Consequently, xylem increases more rapidly in bulk than phloem and soon forms a compact mass. Due to continued formation of the secondary xylem, a pressure is exerted by it on the cambium, phloem and other surrounding tissues. Thus these tissues get pushed outwards. Hence, the primary tissues get crushed. The xylem however remains more or less intact in or around the centre.
Spring and Autumn Wood:
In temperate regions where climatic variations are of pronounced nature, the activity of the cambium is not uniform throughout the year. In spring there is greater production and activity of foliage leaves. Hence, the plant needs more water. Thus, the cambium is more active in spring and forms a greater number of vessels with wider lumen. The wood thus formed is called spring wood or early wood. In autumn, the cambium is less active and forms lesser number of vessels with narrower dimensions. This is known as autumn or late wood.
Spring wood is lighter in colour as their vessels have thinner walls, whereas the autumn wood is darker as their vessels have thicker walls.
These two kinds of wood appear together in a transverse section of the stem, as a concentric ring known as the annual ring or growth ring. Successive annual rings are formed year after year by the activity of the cambium. Each annual ring corresponds to one year of growth, and by counting the total number of annual rings the age of the plant can be approximately determined. The determination of the age of the plant by counting annual rings is called dendrochronology.
Heartwood and Sapwood:
In old trees, the central region of the secondary wood is filled up with tannin and other substances which make it hard and durable. This region is known as heartwood or duramen. It looks black, owing to the presence of tannins, oils, gums, resins etc., in it. The vessels often become plugged with tyloses.
Heartwood does not conduct water but provides mechanical support to the stem. The outer region of the secondary wood which is of lighter colour is known as sapwood or alburnuand this alone is used for conduction of water and minerals.
2. Extrastelar Secondary Growth:
This is the secondary growth taking place in the cortex. The formation of new tissues in the stelar region exerts a pressure on the peripheral tissues of the stem, as a result of which the epidermis gets ruptured. To replace these peripheral tissues a secondary meristem called cork-cambium or phellogen originates from the outer layer of the primary collenchymatous cortex.
This meristematic cork-cambium divides to give new cells on both the sides, forming secondary cortex on the inner side and cork on the outer side. Cells that are cut off on the inner side are parenchymatous in nature and are termed as secondary cortex or phelloderm.
Cells of the secondary cortex generally contain chloroplasts and carry out photosynthesis. New cells cut off by the cork cambium on its outer side are roughly rectangular in shape and soon become suberized. They form the cork or phellem. The three new tissues i.e., phellem, phellogen and phelloderm together constitute the periderm (Fig. 6.27).
The cork tissue of Quercus suber (cork oak) is of sufficient thickness, and is the source of commercial bottle cork. When this is cut off from the plant a fresh strip of cork is produced by the cork – cambium. Being suberized, cork is impervious to water and it cuts off the outer tissues from the supply of water and food material.
Consequently these tissues die and form a part of bark of the plant. All the dead tissues lying outside vascular cambium constitute the bark of the plant. Hence, it includes epidermis, lenticels and cork and sometimes also the hypodermis and a portion of the-cortex, and secondary phloem. It protects the inner tissues and prevents loss of water by evaporation.
Due to the formation of the periderm, the epidermis breaks open forming lenticels. They have characteristic loosely arranged parenchyma cells called complementary cells. Lenticels are aerating pores formed in the bark through which exchange of gases takes place. Externally they appear as small protrusions on the surface of the stem.
A section through a lenticel (Fig 6.27) shows the loose mass of called complementary cells. At each lenticel the cork-cambium instead of producing cork cells, forms oval or irregular loosely arranged cells with inter cellular spaces. The lenticel commonly develops at the place of a stoma and as its cells increase in number and size, the epidermis gets ruptured. Hence, gases from the atmosphere can easily enter in and out through a lenticel. In addition they also help in transpiration.
B. Secondary Growth in a Dicot Root:
Secondary growth in roots occurs in most dicotyledons. It involves formation of secondary vascular tissues in the stele and cortex formed by vascular cambium and cork cambium respectively.
Initiation of Vascular Cambium:
The vascular cambium is formed in the form of strips from the parenchymatous cells present along the inner edges of the primary phloem. The number of the cambial strips depends on the vasculature of the root. Thus, there two strips in a diarch root, three in a triarch root and so on. Later the pericycle cells located outside the xylem also become meristematic and function as the cambium. Now, the vascular cambium becomes a complete and continuous wavy ring running internal to phloem and external to xylem.
Initially, the cambium cells located on the inner face of the phloem are functionally more active than the cambium dedifferentiated by the cells of the pericycle, soon making the wavy cambial ring circular. The cells produced towards the inner-side of the cambium ring so formed differentiate as the secondary xylem while those cut off to the outside differentiate into elements of the secondary phloem. The cambium originating from the pericycle produces wide multiseriate vascular rays.
These rays run radially through secondary xylem and secondary phloem. The primary xylem remains almost in its original position, but the primary phloem is pushed towards the outerside and gets crushed.
Initiation of Cork – Cambium:
The cells of the pericycle undergo periclinal and anticlinal divisions. Thus, a pericycle of many layers is formed.
Increase in secondary vascular tissues combined with increase in the layers of the pericycle ruptures the cortex.
A new cambial ring is differentiated from the outer part of the pericyle. It is called cork – cambium or phellogen. The cells of the cork cambium divide to form the cork or phellem towards the outer side and secondary cortex or phelloderm towards the inside. The pressure caused by formation of secondary tissues ruptures the cortex with epiblema, which is ultimately sloughed off. Here, and there lenticles develop as in the stem.
No comments yet.